RECOMBINACIÓN
La selección natural sólo discrimina fenotipos y éstos son el resultado de la acción cooperativa de todos los genes que constituyen el genotipo. Para que un alelo sea seleccionado, no basta con que sea “bueno” en sí mismo; es necesario además que establezca una armónica combinación con sus “compañeros de viaje”. Si gen muy ventajoso compartiera el genotipo con un alelo letal, también sería eliminado; por el contrario, un gen neutro podría propagarse por el sólo hecho de hallarse ligado a otro favorecido por la selección.
Richard Dawkins, en su libro “El gen egoísta”, establece una analogía entre los genes y un grupo de remeros. Un remero que se vale sólo de sus propios medios no puede ganar una regata. Remar un bote es una empresa cooperativa. El éxito dependerá de qué tan buenos sean los otros miembros del equipo y de la coordinación que exista entre ellos. Siguiendo la analogía de Dawkins, la recombinación equivale a formar equipos de remeros, escogiendo sus integrantes al azar de entre un grupo de ellos (el pool génico). Cabe la posibilidad de que algunos equipos resulten muy malos, pero también es posible que en algún momento surja un equipo que reúna a los mejores.
En las poblaciones, las recombinaciones, ocurridas al azar, son el resultado de la reproducción sexual y tienen lugar en tres instancias:
a. Recombinación gamética: en una población panmíctica, cualquier individuo en edad reproductiva puede cruzarse con otro del sexo opuesto. Esto quiere decir que si en la población hay dos mil individuos en estas condiciones, existen, potencialmente, un millón de cruzamientos (combinaciones) diferentes.
b. Recombinación cromosómica: como se vio en el estudio de la meiosis, la contribución de cromosomas paternos o maternos al complemento haploide de las gametas es un fenómeno aleatorio. Un individuo produce gametas que llevan informaciones sensiblemente diferentes entre sí.
c. Recombinación génica: es consecuencia del crossing-over.
La reproducción sexual, por los mecanismos expuestos, permite la creación de innumerables genotipos nuevos, ofreciendo a las especies el beneficio de una rápida adaptación. En esto reside su ventaja sobre la reproducción asexual, que transmite la combinación genética sin ninguna alteración.
En ciertos casos, sin embargo, la panmixia se ve restringida. Por ejemplo, en las poblaciones pequeñas es frecuente el cruzamiento entre parientes y en las plantas es muy común la autofecundación. Ambas son formas de incesto y limitan la recombinación. El efecto del incesto en una población es la tendencia a la homocigosis (predominio de individuos homocigotas). De esta manera, alelos recesivos que pudieran haberse mantenido ocultos se manifiestan fenotípicamente. Si dichos alelos reducen la posibilidad de adaptación de sus portadores, el incesto resulta nefasto. Pero si, por el contrario, la población ha logrado una combinación genética óptima para las condiciones de su hábitat, la conservación de la condición homocigótica es de vital importancia.
En las poblaciones heterogaméticas o panmícticas se mantiene un porcentaje alto de heterocigosis (predominio de heterocigotas). Esto implica una gran diversidad genética que se recombina en cada generación. Aunque muchas de estas recombinaciones no sean productos o genotipos óptimos, la capacidad de adaptación y la flexibilidad evolutiva alcanzan su máximo.
El genotipo heterocigota es, en muchos casos, más favorable que cualquiera de los homocigotas. A esta superioridad del heterocigota se la llama vigor híbrido o heterosis. No obstante, la población mantiene un polimorfismo balanceado de fenotipos, es decir, los homocigotas no son eliminados, ya que siempre surgen en un 50% de la cruza de dos heterocigotas.
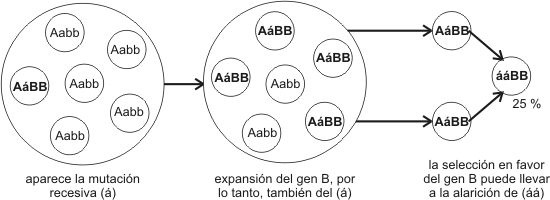
La heterocigosis supone, además, una gran reserva de alelos recesivos ocultos que, por homocigosis, pueden emerger y ser presentados a la selección natural. Suele ocurrir que alelos surgidos por mutación y que permanecen ocultos y neutros por mucho tiempo en un pool génico, emerjan en un momento en que cambian las condiciones ambientales, pasando a ser ventajosos (recordemos el carácter “oportunista“ de la evolución). Este fenómeno se conoce comopreadaptación.